110 years ago, a leading American scientist, Edmund Beecher Wilson (Figure 1), discovered that some specimens of certain insect species, in addition to the species’ normal complement of chromosomes, had some “rare” or “strange” chromosomes. He called these accessory chromosomes, since he could not associate them with any characteristic of the insects when they were present and there were no apparent problems for the insects when they were absent.

Since then, such chromosomes has been found to be present in more than 2,000 eukaryotic species (from fungi and plants to mammals), and the possibility has even been considered of them existing in our own species. But since then, these chromosomes have “resisted” attempts at analysis to clarify their origin and function, if any. In fact, during all this time it was thought they could be parasite or supernumerary chromosomes, or B chromosomes (as referred to from now on) – as opposed to normal As chromosomes. The application of new genetics and molecular biology techniques is beginning to solve these enigmas, and, above all, opens up the possibility of them not being as “parasitical” as we had thought.
Origin
Based on the fact that B chromosomes are usually smaller than those in the normal complement of chromosomes (Figure 2), since their discovery they were considered either to be fragments of a normal chromosome of the species itself, or fragments of a chromosome of a different species that originated following interspecific hybridization processes. At present, although some cases exist (insects like the parasitoid wasp Nasonia or fish like molly Poecilia) where the data indicates that a species’ B chromosomes originate in another different species, in most cases they have an intraspecific origin. In any case, the problem lies in determining from which specific chromosome or chromosomes they originate.

This enigma began to be resolved when the branch of genetics that studies chromosomes, cytogenetics, began to apply molecular techniques (especially techniques of in situ hybridization of DNA) in chromosomes to determine whether certain DNA sequences of the normal chromosomal complement are also present in B chromosomes. In a number of cases, B chromosomes were found to be (mainly) carriers of repeated DNA sequences, or of ribosomal DNA present, exceptionally, in a particular chromosome of the normal complement of chromosomes. It was thus demonstrated that the origin of the B could be associated with a certain chromosome of the normal karyotype of the species in which they exist (1). But nothing could be known about the rest of the sequences that form part of the B chromosomes or whether other different chromosomes were involved.
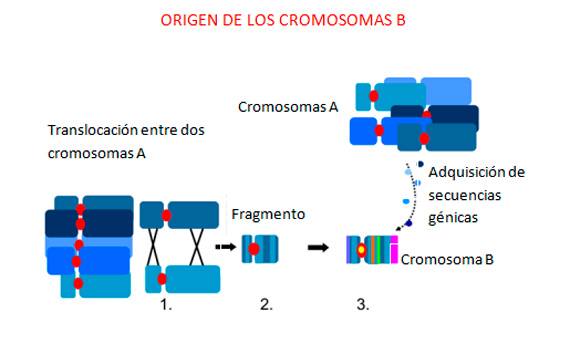
The definitive analysis of the group of genetic sequences that make up B chromosomes was achieved when the new techniques of DNA sequencing began to be applied. Thus, in several animals and plants it has been confirmed that B chromosomes, in addition to repetitive DNA, which in principle is nonfunctional, have functional gene sequences, originating not from just one but from several chromosomes in the normal complement of the species. All of this demonstrates that B chromosomes originate and evolve through complex processes that involve several chromosomes from the normal complement (2) (Figure 3).
Function
Currently, transcriptomics and proteomics are being used to verify how, within B chromosomes, some functional genes are transcribed into RNA and even translated in some cases into added proteins, while in others they regulate the proteins that come from the genes of the normal chromosomes. Thus, organisms with B chromosomes present some additional – and sometimes different and favorable – molecular characteristics compared to organisms that do not have them (3).
For example, it has just been discovered in rye that the presence of B chromosomes determines the overexpression of the activity of the genes that defend plants against thermal shocks, which can be a mechanism of adaptation to heat, which would act mainly in the germinal line and ultimately, in the formation of the grain.
Do they also exist in the human species?
It has also been observed in our species that some people carry in their cells, in addition to the 23 pairs of standard chromosomes, other type B chromosomes, although in this case they are called sSMC (small Supernumerary Marker Chromosomes)(Fig. 4). In some cases, these small chromosomes can be identified as fragments clearly originating from some specific chromosome. However, in other cases it is not clear which chromosomes they come from. Moreover, in most cases such chromosomes have no detectable effect, while in others they do, specifically some clinical symptom. But in almost no case have they been found to be transmitted from generation to generation, as happens with B chromosomes, originating rather de novo in the gametes that have given rise to these people and that most certainly will not be transmitted to the next generation.
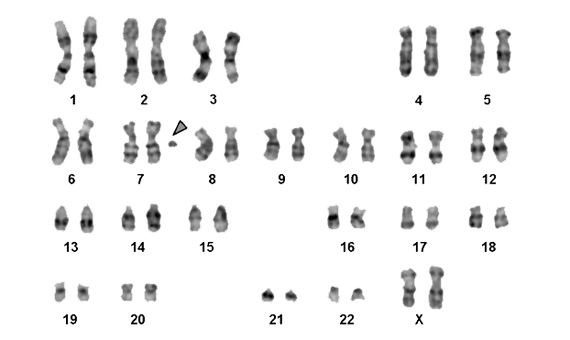
However, a few exceptional cases have been found of people with sSMCs not clearly identified with any particular chromosome, which are not associated with any manifest anomalies, and these have been inherited from their parents and possibly may be transmitted to their offspring. These sSMCs may indeed constitute the true B chromosomes of our species, the presence of which in the general population will have to be determined and studied in the future (4).
It will be interesting to determine whether, as is evident in the Bs of other species, these supposed Bs may contribute in some way to human molecular characteristics. And, in any case, we must pay attention to the incidence of these small sSMC chromosomes and their effects on the human species.
Conclusion
All of the above may mean we will have to change the idea we have had up to now of B chromosomes as non-functional parasites that only take advantage of other chromosomes to replicate and transmit themselves from generation to generation, without contributing anything to the cells and organisms that carry them, and that, in any case – like all parasites –, cause them problems when they accumulate in high numbers.
The case would rather be one of chromosomes originating through complex evolutionary processes from preexisting chromosome sequences, but that then evolve independently, and that may contribute to the functioning and characteristics of cells and organisms, at least at molecular level.
Finally, the last possibility is that certain Bs with functional genes are integrated into the normal karyotype, that is to say that they pass from B to A. This is what is argued to have happened, for example, with the Y sex chromosome of the vinegar fly: it originated as a B chromosome, but later captured the male fertility genes and was integrated in the normal genome of the species.
Manuel Ruíz Rejón
References:
- Ruiz Rejón, M, Ruiz Rejón, C., Oliver, J.L. Evolución de los cromosomas B. Investigación y Ciencia.133: 92-101. 1987; Cabrero, J., Camacho, J.P.M. Los cromosomas parásitos. Investigación y Ciencia. July 2009.
- Houben, A. et al. 2014. Evolution and biology of supernumerary B chromosomes. Cell. Mol. Life Sci. 71:467-478.
- Valente, G.T. et al. 2016. B-chromosomes: from cytogenetics to systems biology. Chromosoma. doi: 10.1007/s00412-016-0613-6.
- Liehr, Th. et al. 2008. Small supernumerary marker chromosomes sSMC in humans: are there B chromosomes hidden among them. Mol. Cytogenet.4; 1:12.; http://ssmc-tl.com/sSMC.html
Comments on this publication