The twentieth century was the century in which human society incorporated technological development in a massive way. For much of that century, the greatest technological contributions grew out of the physical sciences: automobiles, the telephone, airplanes, plastics, computers, and so on. The introduction of those factors has changed society and human behavior more than even political and social events.
During the second half of the twentieth century, however, and especially during the last two decades, biological technology with enormous medical and social potential has emerged. This technology offers a new image of the evolution of life on our planet and is destined to revolutionize the very structure of human society.
Perhaps the person who has most lucidly delved into these ideas is Sydney Brenner. One of the most brilliant scientists of the twentieth century, he will be remembered by the history of science for his enormous contributions to molecular biology, a science he was decisively involved in creating. Brenner says that new biology offers us greater comprehension of ourselves, and a new understanding of humans as organisms: “…for the first time, we can pose the fundamental problem of man and begin to understand our evolution, our history, our culture and our biology as a whole.”
In the present text, I will deal with the history of those scientific events that led to this situation and briefly speculate about the implications these new discoveries have for future society, and even for our own understanding of human nature.
Turning points in biological knowledge
Over the course of its history, biology has undergone three major revolutions. And here we use the term “revolution” to refer to the emergence of a discovery that is important unto itself but also leads to a radical change in the general approach that characterized this discipline until then.
The first revolution took place in 1860 with the evolutionist theories of Darwin and Wallace, who defended the universality of the origin of all living beings. The second revolution was the discovery of the universality of the biological information mechanism proposed by Watson and Crick in 1953. The third revolution has been the discovery of the universality of animal design and that of the basic processes that regulate biological functions. This last revolution took place in the twentieth century, between 1985 and 2000. Unlike the previous ones, it is the result of contributions by a relatively large number of researchers. These three events have led to a new understanding of evolution and of the biology of human beings themselves.
Evolutionary fact
The idea that species change over time is very old and certainly earlier than Darwin’s proposal. In the year 520 B.C., in his treatise, On Nature, Anaximander of Miletus introduced the idea of evolution, stating that life began in the oceans. In his book, Historia Plantarum, published in 1686, John Ray catalogs 18,600 types of plants and proposes the first definition of species based on common descent. And Darwin’s own grandfather, Erasmus Darwin, explicitly proposed that animal species change over time.
What differentiates Darwin and Wallace from their predecessors is that they proposed a plausible mechanism of evolution based on the idea of natural selection. Darwin in particular proposed that the strength of natural selection lay in the survival of the fittest, since their greater capacity for survival also insured them a greater capacity to transmit their characteristics to their progeny. Through this process, the characteristics of populations of each particular species were gradually modified over the course of successive generations.
Darwin also had access to information unknown to his predecessors, and that information contributed considerably to his comprehension of the evolutionary phenomenon. It was known, for example, that the Earth was much older than had previously been thought, which allowed much more time for the gradual change prophesized by the theory of natural selection. By Darwin’s time, there was also a very well cataloged roster of fossils, which made it possible to verify the existence of gradual change in many lines of animals and plants. This clearly supported Darwin’s proposal. It was also known that artificial selection is able to generate very profound morphological changes in a very short period of time. That becomes clear when we consider, for example, the vast variety of breeds of dogs now in existence. They all derive from the wolf, but over the course of five to ten thousand years of artificial—not natural—evolution, man has managed to create a great diversity of canine breeds. This indicates the degree to which biological material is versatile when subjected to selection.
If we were to summarize the implications of evolutionary theory, we would concentrate on three points: 1) all living beings have a shared origin; 2) there has been a process of gradual change over many millions of years that has led to all biological diversity on this planet; and finally, 3) the human species is simply one more of the hundreds of thousands of species that exist or have existed. Darwin’s proposal reflects a Copernican change in the approach to the position of the human species as a biological entity. Man is no longer the center of creation. Instead, he is simply one more species among the millions created by evolution. It is no surprise that there was a great social reaction to this in Darwin’s time. Even now, evolution is not accepted by all members of society. According to the Gallup institute, in 2004, more than half of the United States believed that man was literally created, exactly as the Bible states, some 10,000 years ago.
Genetics and evolution: an operational definition of the gene
Darwin offered a descriptive explanation of biological diversity that was plausible, but not mechanistic. The question is: if all living organisms have a shared origin, what biological function is common to all of them, transmitted from parents to offspring and modifiable in order to generate biological diversity? In his time, Darwin was unable to answer these questions. It was precisely the posing of such questions that led to Genetics, the discipline that studies how biological information is transmitted and modified. We owe the first evidence of the existence of inheritable genetic information to Gregor Mendel, an Augustinian monk who demonstrated that the shape or color of peas is faithfully transmitted from one generation to the next.
But the progress of Genetics in the twentieth century owes much to the fruit fly, Drosophila melanogaster, an organism that has become a classic object of study for genetic research because it breeds easily in laboratory settings, has a very short biological cycle (which is very useful when studying the transmission of diverse traits from one generation to the next) and is totally innocuous to human beings. Drosophila studies revealed many concrete inheritable traits (genes), demonstrating that they are located and aligned in cell nucleae—in organules called chromosomes—and that each gene is situated in a specific position in the chromosome.
They also showed that inheritable variations (mutations) naturally appear in genes, and that these mutations are the source of the biological variation that is essential to the evolutionary process. These mutations can also be artificially induced using radiation or chemical compounds. In sum, what Drosophila genetics discovered is that the real motivating force for evolution are the genes, which make up the inheritable genetic information, and which can be modified.
After over a century of research on this fly, knowledge of its genetics is the most complete of all the animal kingdom, and a series of concepts and technologies have been developed to carry out experiments that are not possible with any other species.
The nature of genetic information
The problem that arose after that, around the nineteen forties, was to discover the gene’s physical nature. What was its chemical composition? The solution to this problem led to what I call the second revolution in biology: Watson and Crick’s explanation of the nature and structure of genetic information as DNA. The famous article published in Nature magazine in 1953 was the beginning of a biological revolution destined to change the very course of humanity. DNA is a molecule with a double-helix structure consisting of two large chains of molecules of a sugar (deoxy-ribose) linked by phosphates. Connecting the two chains, like rungs of a ladder, are other molecules called nitrogenated bases that maintain the structure’s stability. Watson and Crick immediately noticed that the structure of the molecule itself explains the mechanism of replication, leading to identical molecules and thus insuring faithful transmission of biological information for generations.
Moreover, the structure of DNA indicates that biological information lies in the sequence of four nitrogenated bases running throughout the molecule. These bases are called thymine (T), guanine (G), adenine (A), and cytosine (C). What an organism inherits from its progenitors, and which will determine its biological characteristics, is simply a sequence written in a language of four letters.
The discovery of the structure and function of DNA modified biology’s experimental focus: all organisms are encoded in a language of four letters: A, T, C, and G. From then on, biology concentrated on the study of the properties and structure of DNA. The first complete sequence of DNA obtained for an organism, bacteriophage ØX174, contains 5,000 letters (called bases). By comparison, the DNA sequence of a nematode worm consists of 90 million pairs of bases and the sequence of the Drosophila fruit fly contains 120 million pares of bases, while the human sequence has no less than 3,300 million pairs of bases. Each of these sequences represents a sort of formula for the construction of the species in question.
A universal genetic code
The problem is that life processes are not catalyzed by DNA, but instead by proteins. DNA is simply a recipe that has to be translated into the full variety of proteins—some 3,000 basic ones—that control life processes, including the replication and expression of DNA itself.
Proteins consist of combinations of 20 amino acids, so each protein is different from the others because it is made up of a specific sequence of amino acids. Therefore, the sequence of 4 bases inherited from progenitors has to be translated into sequences of 20 amino acids in order to produce the proteins that support biological functions. Deciphering the translation code, the genetic code, was one of the first great successes of molecular biology. The laboratories of Ochoa, Nuremberg, and Brenner were decisive in deciphering the translation mechanism. Those researchers demonstrated that each amino acid is codified by a specific sequence of three bases (triplet), thus insuring that each gene, which is a particular sequence of the complete DNA, is translated into a specific protein. The AAG triplet codifies for the amino acid, lysine, while GCA codifies alanine, and AGA codifies arginine. Thus, the DNA sequence, AAGGCAAGA would translate into the amino-acid sequence lysine-alanine-arginine (see figure 1).
Figure 1. Translation of the genetic text.

What is interesting about the genetic code is that it is universal for all organisms. The universality of this code is, itself, proof of evolution. All organisms have the same genetic code simply because we have inherited it from an ancestral forebear. In this context, a gene is simply a concrete sequence of DNA codified for a specific protein that handles a concrete function, for example, the hemoglobin needed for breathing, or myosine for muscles.
The development of molecular biology
The discovery that DNA is an instruction manual for making a living being, and the deciphering of the basic mechanisms of genetic functions—the genetic code and the manufacturing of proteins—mark the beginnings of molecular biology. Beginning in the nineteen seventies, the study of DNA, its structure, and properties, became the main focus of this discipline. That concentration of efforts has led to extraordinarily powerful concepts that make it possible to manipulate DNA with great efficiency. These are the techniques that allow the cloning of genes, the generation of transgenic animals and plants, the possibility of gene therapy, and the Genome Projects. The generation of transgenic organisms—those in which genes from another species have been inserted—springs from the fact that all DNA, no matter what its origin, is chemically identical, and a gene is simply a fragment of DNA. This makes it possible to use chemical methods to mix fragments of DNA (genes) from different origins. Once methods were developed for inserting those fragments into a receiving organism, that organism could have a gene with a different origin. A clear example of this are strains of yeast into which the human gene that codifies for insulin has been inserted. This procedure has created transgenic yeast that manufactures human insulin.
The great development of these procedures in recent years has made it possible to generate transgenic plants (wheat, soy, rice, and others already on the market) and animals of many species, including rats, mice, pigs, flies, and so on. It is important to note that the methods used for the different animal species are very similar and constitute the basis of applications for their therapeutic use in humans. The goal is to use gene therapy to cure genetic diseases. In 2000, Science magazine published the first test of gene therapy in which several children were cured of a severe immunodeficiency. Unfortunately, those tests had to be interrupted because of harmful side effects. Three of the cured children later developed cancer. This example simultaneously shows both the potential of such new methods and the fact that they are in a very early stage of development. Given the speed with which they are progressing, it is to be hoped that they will be available in the not-too-distant future.
The genetic design of animal bodies
One of the areas in which molecular biology has progressed significantly, and with considerable applications for human biology, is the field of genetic design of animal bodies. Initially, molecular biology experiments used unicellular organisms, bacteria or viruses, to study the properties and functions of DNA. Those studies produced very important results, as described above, but their very nature made it impossible to draw conclusions about genetic control of the development of complex organisms, such as a fly or a mouse, in which associations of cells have to be grouped in the proper fashion as part of a three-dimensional structure.
Let us consider, for example, a butterfly (figure 2). Each individual cell has to carry out the primary biological functions—protein synthesis, replication of DNA, and so on—but it must also be able to form groups with other cells and differentiate itself in order to make specific organs such as eyes, wings, legs, and so on. Those organs have to be assembled with the other organs in order for each to appear in the right place. An animal design calls for the various parts of the body to be properly situated in space’s three dimensions: the anterior-posterior, dorsal-ventral, and proximo-distal axes. This problem of body design has been one of the great challenges to the genetics of superior organisms: how genes specify positional information for different parts of the body so that the cells that are going to make an eye know they have to do so in the upper part of the body, and those that make the legs have to be in the ventral part. In other words, what is the genetic description of a three-dimensional organism? In an insect like a butterfly, we can morphologically distinguish a cephalic part, a thoracic part and an abdominal part, but there is no guarantee that this description corresponds to the true genetic description of the organism.
There has been notable progress on the question of the genetic description of animals in the last thirty years. The keys to its genetic design are in what are called homeotic genes, now called Hox. These make up genetic mechanisms that have been studied in great detail in the Drosophila fruit fly. What is characteristic of these genes is that their mutations transform some parts of the body into others (figure 3). A mutation such as Antennapedia (Antp), for example, transforms an antenna into a leg, while a mutation such as Ultrabithorax (Ubx) transforms the halter into a wing, creating a fly with four wings. What is interesting about these transformations is that, despite the fact that the general construction of the body is erroneous, the morphology of the parts is normal: the leg that appears in the antenna of Antp is normal, only its location is anomalous. Likewise, the transformed wings that appear in Ubx flies have the normal size and shape of wings. The only abnormality is where they appear. The implication of those phenotypes is that what Hox genes control is not the morphology of the structures, but rather the general design of the body, the positional information I mentioned above, which insures that each organ appears in its proper place.
Homeotic genes are thus high-level regulator genes that determine the type of development of the different parts of Drosophila’s body. A very important question that arose in the nineteen eighties was: how many homeotic genes exist? It was hoped that the identification of all of them would make it possible to clarify the genetic logic underlying the body’s design. Studies carried out in the United States and in Spain demonstrated that the number of Hox genes is surprisingly small. There are only nine such genes in Drosophila, where they establish the spatial coordinates of the anterior-posterior axis, recognize the positional value along each axis, and determine the acquisition of the proper development program for generating each part of the body. These results were certainly interesting, but they referred to the fruit fly. At first, we did not suspect that they might have a general value in explaining the body design of other animals, including the human species.
Nevertheless, the progress of molecular biology in the nineteen seventies and eighties permitted the molecular isolation (cloning) and sequencing of Drosophila’s Hox genes. By late 1985, all of those genes had already been cloned and sequenced. An extraordinarily important discovery was made when their sequences were compared: they all had one sequence in common, which was named homeobox. The discovery of the homeobox sequence had very important implications: 1) this sequence codifies for a motif of union to DNA, indicating that homeotic proteins function as transcription factors and regulate the activity of other subsidiary genes; 2) the presence of the same sequence in all Hox genes indicates that those genes have a shared origin; and 3) the homeobox sequence is a molecular marker for Hox genes that makes it possible to identify these genes in organisms (the human species, for example) in which it is impossible to detect them with conventional genetic procedures. As we will see below, this last aspect proved very significant.
A universal genetic design
The fact that the homeobox is a molecular marker in Hox genes made it possible to identify the Hox complex in many groups in the animal kingdom, making these genes a fundamental subject of biological research in the nineteen eighties and early nineties. The general result is that the Hox complex has been found in all animal groups in which it was sought. It is thus a universal characteristic of the genome of all animals, including the human species. Humans have a Hox complex that is very similar to that of Drosophila, except that, instead of having one copy in each genome, we have four.
Drosophila studies had previously established that the function of those genes was to determine the development of the different parts of the body, but there was no evidence as to their function in other organisms. The difficulty of studying this aspect is that the genetic analyses carried out on Drosophila are not possible
in many vertebrates and totally impossible in the human species. Therefore, other methods had to be employed.
The molecular technologies developed in the nineteen eighties and nineties made it possible to generate individuals—in this case, Drosophila fruit flies—into which the gene of another species could be inserted in order to study its function in that foreign system. Various experiments of this kind made it possible to conclude that the Hox genes of humans and other vertebrates work in identical or similar ways to those in Drosophila. The conservation of functions is such that human or mouse genes are able to replace their counterparts in Drosophila. That is the case of mouse gene, Hoxd13. When inserted into the fly, it is just as capable of programming the development of the rear part of Drosophila as the fly’s own gene. Other very striking examples are, for example, genes of apterous and eyeless Drosophila, which have known counterparts in humans. Apterous is needed to make wings and its mutations produce individuals without wings. Eyeless is needed to program the development of the eye, and individuals in which this gene has mutated do not have eyes.
When a mutant apterous fly receives the human gene, it is capable of forming fly wings. Thus, even though humans do not have fly wings, we have a gene capable of replacing the Drosophila gene that programs the formation of fly wings, thanks to a mechanism of functional conservation. In that same sense, the mouse gene that is homologous to eyeless, called small eye, is capable of inducing fly eyes (figure 4). Similar experiments with genes from other organisms have led to the conclusion that the genetic design of eyes is the same in all animals, be they flies, octopi or human beings. The evolutionary invention of a light-receiving organ connected to the brain took place around 540 million years ago and has been inherited by all multi-cellular organisms. These experiments illustrate a general principle of the phenomenon of evolution: when a mechanism that works adequately appears, the genetic programming of that mechanism remains fixed in the genome and stays the same, or with only slight modifications, from then on.
The general conclusion of all the above is that the overall mechanism of genetic design of animals, based on Hox genes and their derivatives, is common throughout the animal kingdom.
The explosion during the Cambrian era, that is, the sudden apparition of bilateralia with organs arranged along all three of the spatial axes, is almost certainly the result of the apparition of the Hox complex and its derivatives during the lower Cambrian. The similarity of sequences among these genes indicates they come from an ancestral gene that underwent various duplications in tandem, thus generating the set of linked genes that make up this complex. We can thus state that all living beings share the same basic biological functions. Together, these studies have given rise to a unifying view of biological processes based, ultimately, on the evolutionary process. As Darwin and Wallace proposed, organisms have a common origin, sharing the mechanism that stores and releases genetic information based on the universality of the function of DNA, RNA, and the mechanism of genetic code. Finally, all members of the animal kingdom share the same genetic process of body design.
An important implication of these observations is that many aspects of the human body’s design can be studied in model organisms such as flies, worms, or mice. It is understood that the genetic/molecular base of those processes is common to all species, and therefore, many of the processes involved will be so as well. A typical example of this approach can be found in regeneration studies being carried out on amphibians and chickens. It has long been known that amphibians and reptiles are able to regenerate their limbs, while birds and mammals cannot. The studies underway are making it possible to identify genes related to the regenerative process, several of which are also present in species that do not regenerate. It seems that the capacity to regenerate an organ or not depends less on the presence or absence of one or more genes than on the mechanism that regulates common genes. Regenerating species are able to activate these genes following physical trauma while non-regenerators are not. A well-founded speculation is that when the process that regulates those genes is understood, it will be possible to intervene in the control of its functioning in order to artificially induce the regenerative process in species like the human one, which cannot do it naturally.
The genome projects
What has been set out above is, itself, proof of the entire phenomenon of evolution, as it clearly shows the functional universality of biological phenomena. Furthermore, new molecular technology has offered us a more direct demonstration of this universality. In recent years, the complete sequences of DNA (the genome projects) for many animal and plant species have been completed, making it possible to directly compare the degrees of similarity or difference in the biological information of different species.
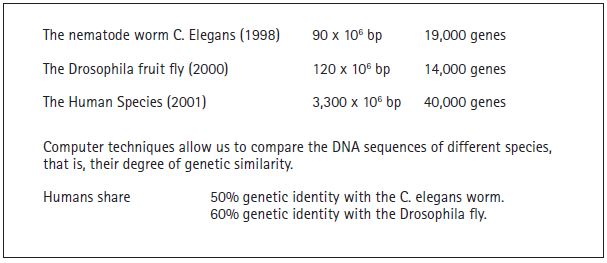
Particularly relevant in that context are the genomes of the nematode Caenorabditis elegans, which contains DNA with 90 million pairs of bases; of the Drosophila fly, with 120 million pairs of bases; and of the human species, with 3,300 million pairs of bases. The Human Genome Project (figure 5) used the DNA of five people (three women and two men) from four different ethnic groups (Hispanic, Asian, Afro-American, and Caucasian). It is interesting to note that no significant differences were detected among them. These projects have managed to identify all the genes in each species, determining their sequence and accumulating that information in databases. Along with the development of very sophisticated software and powerful computers, this has made it possible to compare the significant sequences. That comparison has produced many interesting results, one of the most important of which (figure 6) is the discovery that the human species shares approximately 50% of its genes with those of the nematode Caenorabditis elegans and about 60% with the Drosophila fruit fly. This observation is a healthy reminder of our biological origins, which we share with the rest of the animals. Naturally, this is reflected in the DNA that is the common evolutionary record linking us all.
Figure 6. A comparison of some important genomes.
The study of human illness in model organisms
The high degree of genetic similarity among the species mentioned and, in fact, throughout the animal kingdom, not only validates the phenomenon of evolution; it also has powerful implications for the study of human biology and pathology. Because we share so many genes with organisms such as Drosophila, there are many aspects of biology and human illness that can be studied in flies without the experimental and ethical limitations imposed by human material. The philosophy underlying this is that much of the knowledge obtained by working with Drosophila will also be applicable to us. As we saw above, the study of Hox genes of flies is shedding very important light on the function of those same genes in our own species.
With regard to pathological processes, the latest estimates indicate that 75 percent of genes related with human illness are present in Drosophila. That makes it an enormously important source of information for basic knowledge of human illness. Currently, numerous laboratories around the world are using Drosophila as an organism for studying pathologies such as cancer, Alzheimer’s disease, ataxias, and so on. One example of this approach is the experiments that seek to induce the molecular syndrome of Alzheimer in Drosophila. Deposits of the protein, amyloid (Aß), in neurons is a characteristic of that illness. The pathological form contains 42 amino acids rather than 40 and forms aggregates called amyloid plaques. Drosophila technology makes it possible to induce this illness in the eyes and brains of a fly and to study its evolution. It is possible to produce hundreds of individuals and test a large number of possible remedies or compounds that interfere with the development of the illness. Those experiments have made it possible to identify a drug (Congo Red) that considerably mitigates the effect of this illness in flies. Although the drug is toxic for humans and cannot be used to treat the illness, it clearly indicates the potential of this type of technology. Experiments of this kind have already identified various drugs aimed at treating cancer and other degenerative processes.
Can the duration of human life be changed?
The extremely high degree of conservation of fundamental biological phenomena throughout the animal kingdom allows us to speculate on the possibility of manipulating processes only recently considered inaccessible to human intervention. One of the fundamental paradigms of human society and culture is the idea that aging and death are inevitable biological processes. The supposition is that there is internal programming that establishes the maximum lifespan of members of each species within a relatively narrow range.
During the twentieth century, the average human lifespan increased considerably, due mainly to improved living conditions, hygiene, and medical progress. Even so, the estimated maximum lifespan is about 120-125 years. Could this limit be surpassed? That is a subject that has received considerable attention in international science magazines (Nature 458, 2008, 1065-1071), fundamentally because of recent discoveries directly related to the genetic programming of lifespan.
The fundamental fact is that, in both the nematode worm, Caenorhabditis elegans, and in the Drosophila fly, various genes have been identified whose function is directly related to the aging program of those species. Given the ease with which those organisms can be genetically manipulated, it has been possible to substantially prolong the life of individuals from those species. In the case of nematodes, lifespan has been successfully multiplied by six- or even seven-fold. If this were extrapolated to the human species, it would offer an average human lifespan of some 350 years, and some individuals would live over half a millennium.
What is important about these discoveries is that the aging genes identified in the nematode worm and in Drosophila are also present in the human species. The most studied of those genes, called DAF-16 in worms and FOXO in Drosophila and humans, is related to the insulin path and some of the variant forms of FOXO appear to be particularly frequent in individuals over one hundred years old. Mutations in the human species that affect the activity of the insulin path have also been detected in individuals who are over one hundred. DAF-16/FOXO has been cloned and genetically modified worms have been created in which alterations in the levels of this gene’s functions result in alterations that double the lifespan of those worms. The fact that such results can be obtained by altering just one gene illustrates the potential of such techniques. As we mentioned above, this gene is present in our own species, which suggests the possibility that its manipulation could be used to modify the lifespan of human beings.
The future evolution of the human species: technological man
In closing, I would like to briefly reflect upon the evolution of life on our planet, and the life of the human species. Life on our planet began around 2,000 to 3,000 million years ago. Bilateralia animals, the animals that exist today, appeared around 540 million years ago. Around 100,000 to 200,000 years ago, Darwinian selection led to the human species, along with many millions of others, living or extinct. However, the intellectual and technological development of our species has made it especially immune to the process of natural selection. As a result, normal rules of evolution have little or no effect on us nowadays.
Human civilization began some 10,000 years ago and technological development about 200 years ago. DNA technology is about 25 years old. This technology has progressed extremely rapidly, leading to very powerful methods of manipulation. In sum, the vehicle of evolution, DNA, is being modified directly by human intervention. These methods, though still very crude, are being used on experimental animals—flies, mice, worms, and so on—whose great genetic similarity to us indicates that the day is not far off when they can be applied to the human species. These methods have enormous potential, especially when we consider that they only began twenty-five years ago. It is impossible to imagine what they will be able to achieve in another fifty years, not to mention 500 or 5,000. The human species will be able to genetically modify itself in a controlled manner. That perspective offers enormous possibilities for determining our own biological future and evolution. DNA technology offers a new social paradigm, and will be able to completely change the very essence of the human being.
Comments on this publication